主龙类和龟类的蛋壳钙质层由壳单元组成,其中从壳膜上生长出来的称为初生壳单元,从钙质层内部生长出来的称为次生壳单元。尽管在现代鸟蛋中非常罕见,次生壳单元在恐龙蛋中却是一个常见的结构。由于对这种结构缺乏深入的研究,学界对它是生物成因还是非生物成因仍有争议。产自中国的多种恐龙蛋壳都具有次生壳单元,我国的学者普遍认为它是生物成因,并且可以作为恐龙蛋的分类特征使用;欧洲的学者基于对产自当地的蜥脚类恐龙蛋壳的研究,认为它是非生物成因。
近日,中国科学院古脊椎动物与古人类研究所的高级工程师张蜀康、博士后Seung Choi(现首尔大学研究教授)与浙江自然博物院、韩国首尔大学、荷兰乌特勒支大学和西班牙巴塞罗那自治大学的科研人员合作,综合运用电子背散射衍射(EBSD)、偏光显微镜(PLM)、扫描电镜(SEM)和透射电镜(TEM)等技术,揭示了恐龙蛋次生壳单元的成因,并对其功能、生长方式和演化模式进行讨论。论文发表在Science Advances上,张蜀康为第一和通讯作者,Seung Choi为第二作者。
该项研究分析了恐龙蛋的大部分主要类群,并且用现生鸟蛋、龟蛋和鳄鱼蛋作为对比材料。EBSD分析结果显示,恐龙蛋大部分次生壳单元的c轴都围绕生长核心呈放射状排列,并且平行于蛋壳生长方向延伸。只有少数靠近气孔道的次生壳单元,其c轴向气孔道方向延伸。另外,这些次生壳单元在晶界图上具有带纹理的灰色背景,并且显示出较高的KAM值。
恐龙蛋次生壳单元的上述结晶学特征与其初生壳单元几乎完全一致,更重要的是,也与现生龟蛋和鳄鱼蛋的次生壳单元的结晶学特征一致,这表明恐龙蛋的次生壳单元应当为生物成因的结构。SEM和TEM的分析显示,恐龙蛋的次生壳单元具有许多细小的槽和孔洞,与鸟蛋的壳单元很相似。这些槽和孔洞是基质纤维在埋藏过程中降解后留下的空间。值得注意的是,一些传统的判断恐龙蛋壳中生物成因与非生物成因方解石的方法,如阴极发光(CL),不能很好地区分这两种方解石。因为方解石是否发出橙色荧光与铁离子和锰离子的含量相关。
在一些气孔道发达的恐龙蛋壳里,次生壳单元重叠在初生壳单元之上,或者在气孔道内生长。这些次生壳单元在形态和大小上都与初生壳单元相似,也具有与初生壳单元相似的功能,即作为蛋壳的结构成分,对胚胎起保护作用;在气孔道内的次生壳单元还可以降低蛋壳的气体传导率,防止胚胎在发育过程中过度失水。在一些气孔道较少的恐龙蛋壳里,次生壳单元常常被包裹在初生壳单元或其他次生壳单元内。这些次生壳单元没有明确的功能,或者说它们的功能已被包裹它们的壳单元取代。值得注意的是,在气孔道发达的恐龙蛋壳里,那些生长过程不受初生壳单元和其他次生壳单元干扰的次生壳单元的c轴也平行于蛋壳生长方向延伸,这个现象挑战了基于对鸟蛋壳的研究而提出的“竞争假说”,即壳单元的c轴方向是由基质纤维控制的,而不是相邻壳单元的方解石晶体在生长过程中相互竞争的结果。
早期的研究曾经提出,具有次生壳单元的恐龙蛋壳的与现代的楔齿蜥(Sphenodon punctatus)蛋壳相似,壳单元与壳膜纤维同时生长,而不像现代鸟蛋、龟蛋和鳄鱼蛋那样先产生壳膜,再形成壳单元。本次研究则显示,具有次生壳单元的恐龙蛋壳的生长方式与现代龟蛋和鳄鱼蛋相同,壳膜先形成,然后基质纤维与壳单元同时生长,次生壳单元的形成是由基质纤维而不是壳膜纤维控制的。
尽管大多数恐龙蛋与产蛋恐龙不能对应,但本次研究涵盖了蜥脚类、鸭嘴龙类和可能的基干坚尾龙类产的蛋。这些恐龙的蛋壳都具有次生壳单元,而包括鸟类在内的手盗龙类的蛋壳则极少出现次生壳单元。这说明兽脚类恐龙在向鸟类演化的过程中,蛋壳的形成机制发生了变化。手盗龙类蛋壳的基质纤维与其他恐龙不同,能够产生更加有序的亚层结构,而不会相对随机地产生次生壳单元。另一方面,次生壳单元在龟类、鳄类及鸟臀类、蜥脚类和兽脚类恐龙支系中都有出现,而这些支系的蛋壳钙质层可能是各自独立演化出来的。这说明它们的次生壳单元可能是随着其蛋壳钙质层的演化而独立演化出来的。不过,从生物矿化的分子机制的层面上来说,也不能排除上述所有支系的次生壳单元具有深层同源性的可能。
该研究得到国家自然科学基金(42272020)的资助。
论文链接:https://doi.org/10.1126/sciadv.adt1879
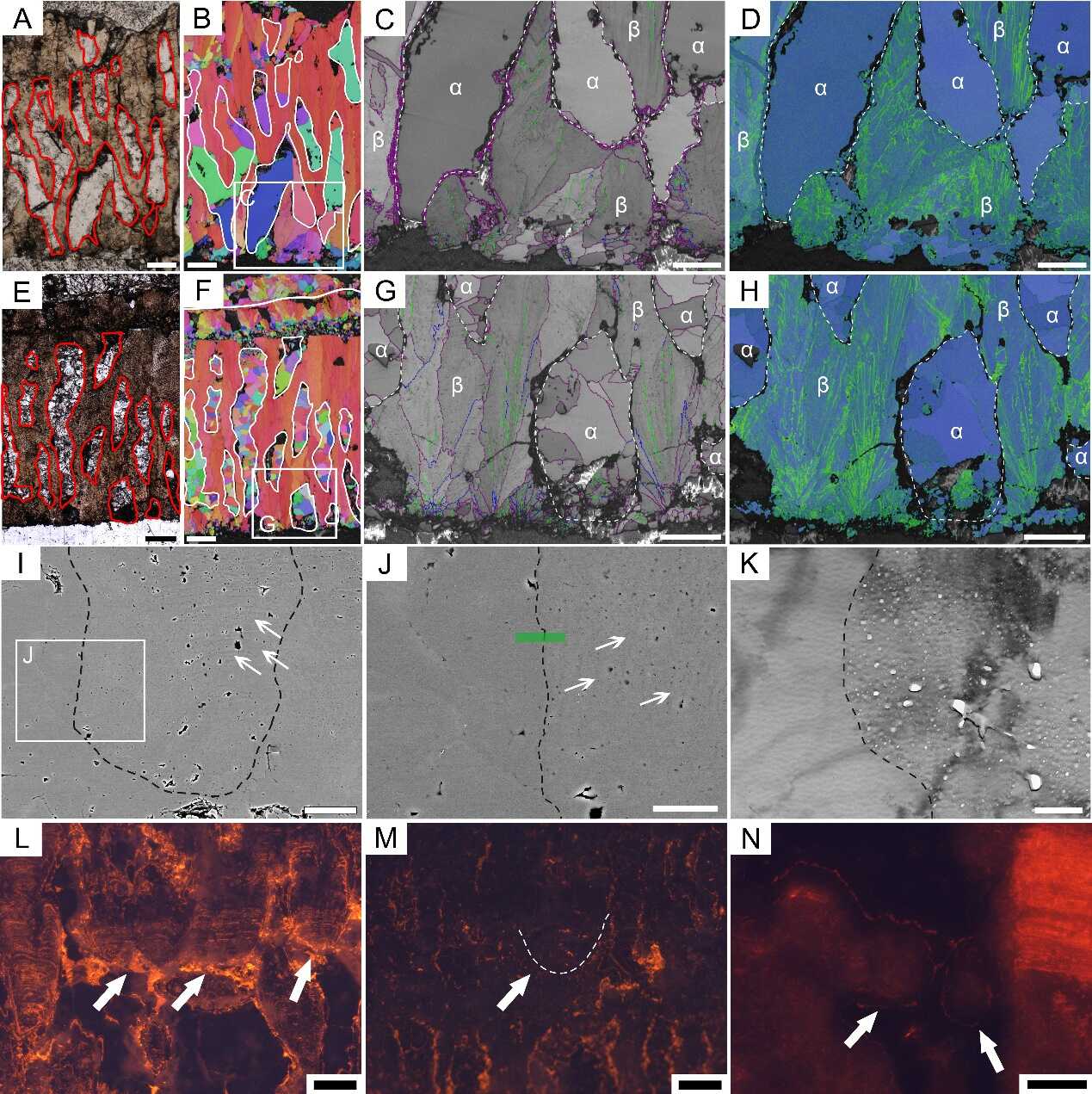
恐龙蛋壳内生物成因和非生物成因方解石的区别:PLM图像上(A和E),生物成因方解石为暗色,非生物成因方解石为亮色;反极图上(B和F),生物成因方解石c轴方向比较一致(红色),非生物成因方解石没有固定的c轴方向(多种颜色);晶界图上(C和G),生物成因方解石具有带纹理的灰色背景,非生物成因方解石的背景比较干净;KAM图上(D和H),生物成因方解石有较高的KAM 值(绿色),非生物成因方解石的KAM 值较低(蓝色)。SEM图像(I和J)和TEM图像(K)显示生物成因方解石具有生长纹和微小的孔洞(箭头),非生物成因的方解石则没有。CL图像上(L—N),次生壳单元(箭头)既可以显示橙色荧光,也可以不显示橙色荧光,因此不能用这种方法判断次生壳单元的成因。比例尺=200 μm (A、B、E和F),100 μm (C、D、G、H和N),50 μm(I),15 μm(J),500 nm(K)和150 μm (L 和 M)。
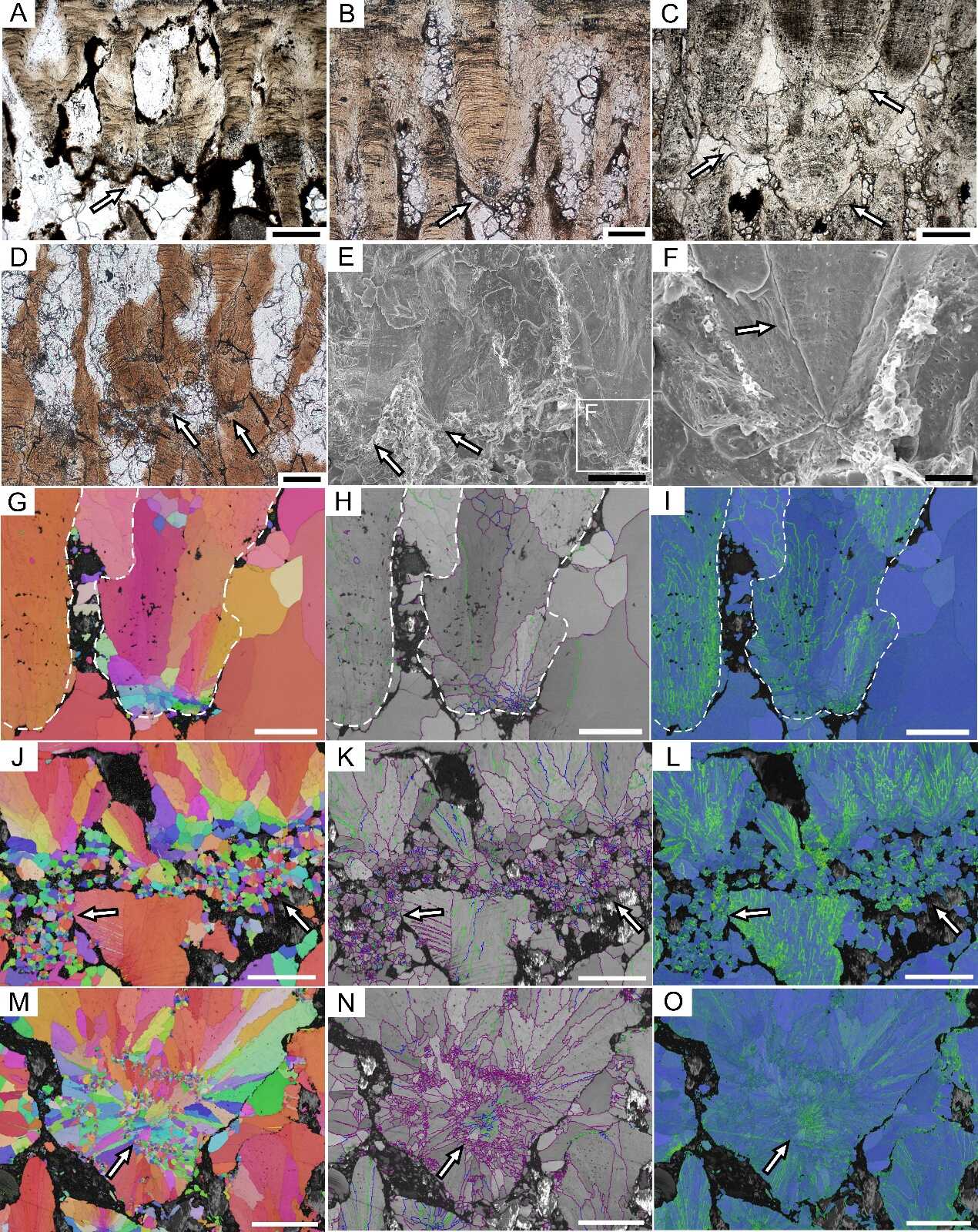
多气孔的恐龙蛋壳内的次生壳单元。A—D:PLM图像;E、F:SEM图像;G—O:EBSD图像。次生壳单元的c轴都围绕生长核心呈放射状排列(G、J和M),在晶界图中具有带纹理的灰色背景(H、K和N),并且显示出较高的KAM值(I、L和O)。F中的箭头示次生壳单元内基质纤维降解后留下的沟槽和孔洞,其他图像中的箭头示次生壳单元。比例尺= 200 μm(A 和C),100 μm(B、D、E和G—O)和25 μm(F)。
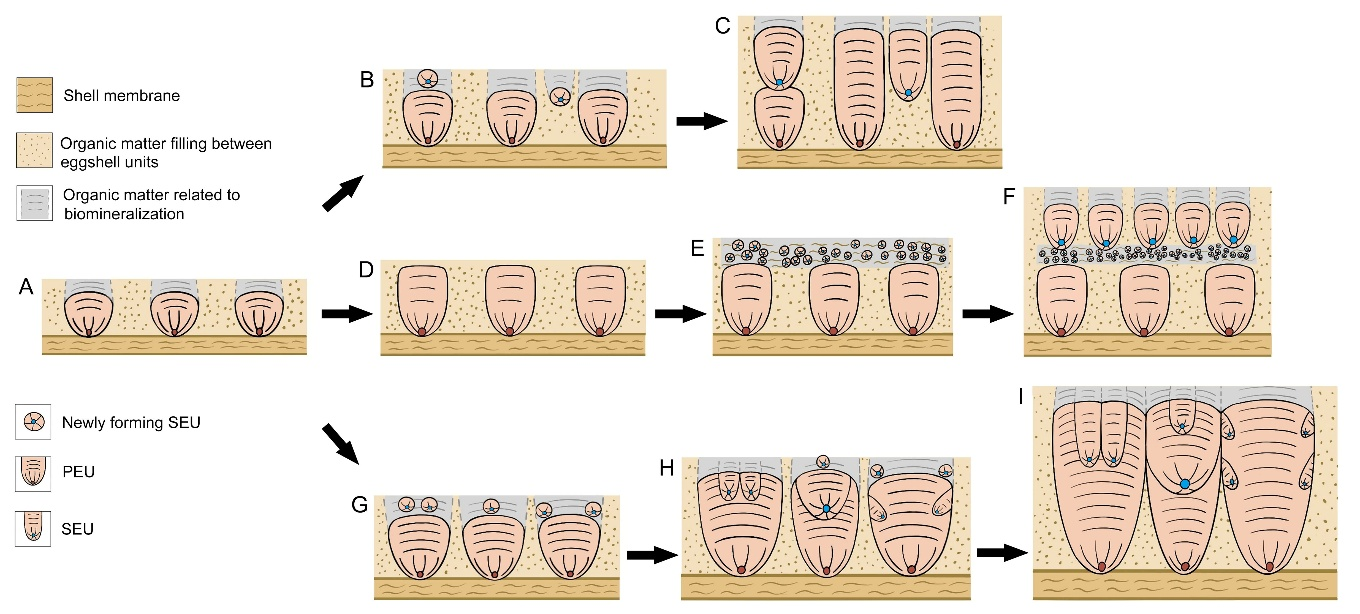
次生壳单元的生长方式。B—F:多气孔的恐龙蛋壳,次生壳单元重叠在初生壳单元之上,或生长在气孔道内(B和C),或形成层状结构(E和F);G—I:气孔较少的恐龙蛋壳,次生壳单元多被初生壳单元或其他次生壳单元包裹。
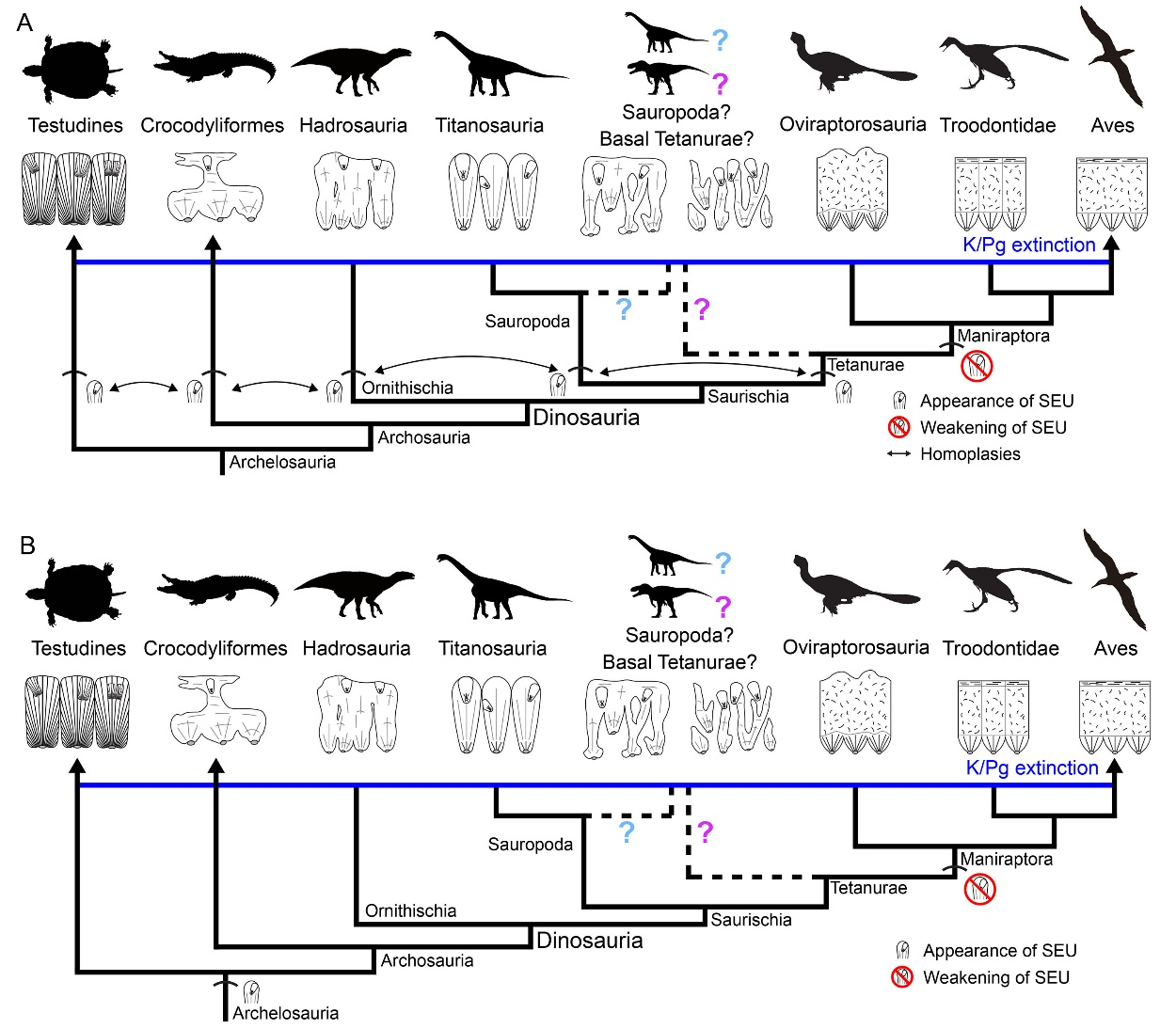
次生壳单元在手盗龙类蛋壳中逐渐消失。龟类、鳄类、鸟臀类恐龙、蜥脚类恐龙和兽脚类恐龙的次生壳单元可能是平行演化的结果(A),也可能具有深层同源性(B)。